Obesity is a major issue in developed countries. Regulating thyroid hormones is a growingly popular adjunct in a comprehensive approach towards weight loss. Despite growing popularity, there is a lack of knowledge about underlying pathways. This review investigates the thermogenic properties of thyroid hormone on brown adipose tissue (BAT), white adipose tissue (WAT) and the hypothalamus. Current evidence about potential inducers of thermogenesis and potential clinical implications for weight loss are explored.
Thyroid hormone production is tightly regulated and clinical manifestations of thyroid system dysfunction are well known. Production of thyroid hormone is maintained by negative feedback loops in the anterior pituitary and the hypothalamus (Figure 1). This represents an important homeostatic center, known as the hypothalamus-pituitary-thyroid (HPT) axis.1 The thyroid secretes iodothyronines under stimulation of thyroid releasing hormone (TRH) and thyroid stimulating hormone (TSH). Thyroid hormones include 3′,5′,3,5-L-tetraiodothyronine (T4), 3′,5,3-L-triiodothyronine (T3), 3′,5′,3-L-triiodothyronine (rT3), and 3,5-L-diiodothyronine (T2). T4 is the dominant iodothyronine secreted by the thyroid and it is regulated by various intracellular pathways.2 Notably, 5′-deiodinase type 2 (D2) is an important enzyme found in the hypothalamus, white adipose tissue (WAT), and brown adipose tissue (BAT). It is essential for converting T4 to the more biologically active form T3.3 The interaction of T3 with its nuclear receptors in target cells results in increased rate of DNA transcription, which induces changes in mRNA, protein, and cellular function.2,4 One of the various physical manifestations of these molecular changes is the production of heat. Thyroid hormones play an integral role in thermogenesis and in maintaining body temperature. Indeed, core body temperature is often regarded as a proxy for metabolic status. As such, maintaining an optimal basal metabolic rate is strongly influenced by thyroid health. In this review, we discuss the thermogenic properties of thyroid hormones and how their use may be ideal to improve the lives of patients with various chronic illnesses, such as obesity and metabolic syndrome.
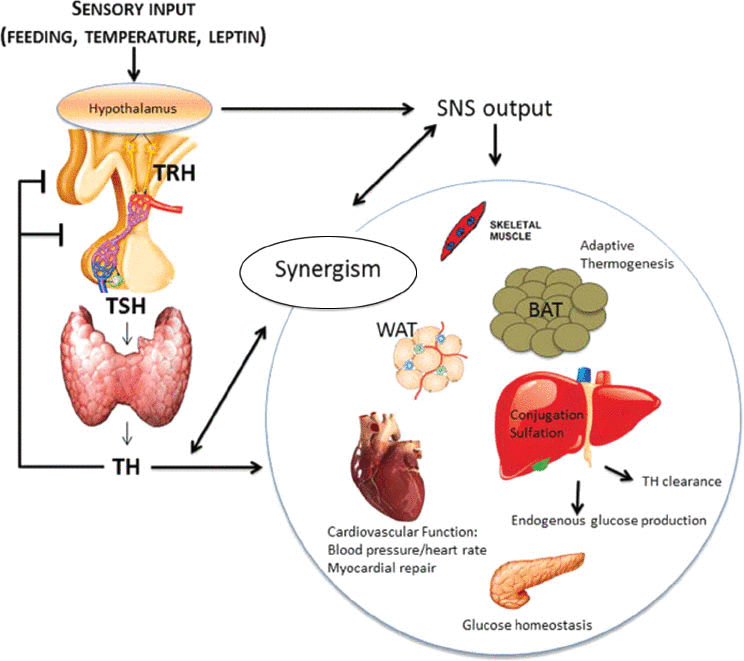
Figure 1:The role of thyroid hormone in homeostasis.
TRH, thyrotropin-releasing hormone; TSH, thyroid stimulating hormone; TH, thyroid hormones; SNS, sympathetic nervous system; WAT, white adipose tissue; BAT, brown adipose tissue.
(From McAninch EA, Bianco AC. Thyroid hormone signaling in energy homeostasis and energy metabolism. Ann NY Acad Sci. 2014;1311:77–87.)
The subject of this review is the thermogenic mechanisms of thyroid hormones. Thyroid hormones play an essential role in homeothermia. Hypofunction or hyperfunction of thyroid hormones is associated with clinically significant cold and heat intolerances.5 Homeothermic species have basal and adaptive mechanisms to ensure a normal range of core body temperature. Since they often live in external temperature environments that are non-thermoneutral. Thermoneutrality is defined as the range of environmental temperatures in which basal thermogenesis is adequate in maintaining core body temperature without the need to recruit any adaptive mechanisms.5 Various evolutionary thermogenic mechanisms exist to create heat beyond our basal metabolic rate and are critical in maintaining core body temperature (TC).5,6 Thermogenic mechanisms are under both thyroid and neural control; the activation of these is dependent on the discrepancy between ambient temperature and core body temperature.
THYROID HORMONE AND BASAL THERMOGENESIS
Basal thermogenesis is defined as the heat created irrespective of ambient temperature, and is also known as basal metabolic rate. Basal thermogenesis is a byproduct of life-sustaining biochemical reactions. There is an innate inefficiency with the utilization of energy, notably adenosine triphosphate (ATP), that results in energy loss as heat.
THYROID HORMONE AND ADAPTIVE THERMOGENESIS
Adaptive thermogenesis is created on demand to maintain TC and allows for greater capacity to live in variable temperature environments. Both shivering and non-shivering mechanisms comprise adaptive thermogenesis. Non-shivering thermogenic mechanisms are more energy dependent but do not impair locomotion. They are controlled by the central nervous system (CNS), when cold-sensitive thermoreceptors are stimulated and norepinephrine is released.7 Adaptive thermogenesis represents the metabolic rate change in response to environment, nutritional modifications, and body composition. It includes activity-related energy expenditure (AEE), diet-induced thermogenesis, cold-induced thermogenesis, and resting energy expenditure (REE). Adaptive thermogenesis, most notably REE in humans, is influenced by thyroid hormones, leptin, and the sympathetic nervous system (SNS). Nutritional starvation results in reduced T3 by way of peripheral thyroid metabolism and the HPT axis.1,4,8 Thyroid thermogenesis is important in the following tissues: BAT, WAT, and the hypothalamus.9
THYROID THERMOGENESIS AND BROWN ADIPOSE TISSUE
Until recently, the presence of BAT had been established in humans up until infancy and was long accepted to be absent in adults.10–14 In 2009, this hypothesis was challenged when F-fluorodeoxyglucose (18F-FDG) positron-emission tomographic (n=2640) and computed tomographic (PET-CT) scans (n=1972) were evaluated for the presence of BAT. The results demonstrated that women were more than two times likely to have detectable BAT than men (7.5% vs. 3.1%, respectively). It was also found that the presence of BAT is inversely proportionate to age, body-mass index and ambient temperatures.15
Thermogenesis in BAT is mediated by uncoupling protein (UCP)-1. UCP-1 is responsible for the uncoupling of oxidative phosphorylation resulting in a proton leak across the inner membrane of the mitochondria. The sequential signaling cascade leads to inefficient ATP production and energy being lost as heat.5,7,16–19 T3 receptor β1 isoform modulates the expression of UCP-1 and D2 in BAT and thus is an important activator of energy expenditure. In a study by Enerbäck et al., UCP-1 deficient mice had impaired thermoregulation and were cold-sensitive. Contrary to expectations, UCP-1 deficient mice did not gain weight with a hypercaloric diet.20 It was discovered that UCP-2, a homologue of UCP-1, is overexpressed in UCP-1-deficient mice compared to controls. UCP-2 overexpression in the setting of UCP-1 deficiency counteracts the losses in energy from various tissues. These data suggest that impaired thermogenesis is not a causal factor of obesity or weight gain.20
T3 also plays an important role in regulating adaptive thermogenesis through the binding of thyroid receptor-α (TRα). The results of a study evaluating homeothermia in thyroid receptor-α knockout (TRα-0/0) mice demonstrated that the animals had cold intolerance due to defective adaptive thermogenesis in BAT and also exhibited central dysfunction at the level of the hypothalamus.18
More recently, studies indicate that ambient temperature can influence metabolic parameters and susceptibility to weight gain in D2 knockout mice (D2KO). D2KO mice have impaired D2 and are therefore exhibiting a hypothyroid state. At environmental temperatures below that of thermoneutrality, also known as hypothermoneutrality, D2KO hypothyroid mice do not show an impaired metabolic phenotype. Only at thermoneutrality do D2KO hypothyroid mice demonstrate significantly slower metabolic expenditure than that of control mice. At hypothermoneutrality, mice down-regulate muscle thermogenesis and caloric intake, while maintaining thermogenesis in BAT. Only at thermoneutrality do mice gain weight on a high-fat diet.21
Emerging evidence suggests that BAT transplantation in mice on a high-fat diet can improve whole body energy regulation as well as metabolic parameters such as insulin sensitivity and glycemic control.22,23 Liu et al. explored if the same benefit could also be conferred in genetically susceptible obesogenic mice and leptin-deficient ob/ob mice. BAT from C57B/L6 mice was transplanted into leptin deficient ob/ob mice. The ob/ob mice had significantly lower weight gain, fat mass, liver steatosis, and insulin resistance.1,24 Hormonally, BAT transplantation reduced levels of free T3 and T4 and increased levels of adiponectin, a major hormone secreted by adipose tissue, which is important in fatty acid oxidation and insulin sensitivity.3,4,25,26
In humans, the effect of thyroid hormones on the post-prandial thermogenic effect of glucose in hypothyroid and euthyroid women was evaluated. Energy expenditure was assessed at rest and throughout oral glucose tolerance testing.27 Significant elevations of catecholamine, glucose, and insulin were observed in the hypothyroid cohort. These results support the notion that hypothyroid subjects have a decreased thermogenic effect of glucose as well as poorer insulin and catecholamine sensitivity compared with their euthyroid counterparts.27
A study by Zhang et al. explored BAT in hyperthyroid patients with Graves’ disease via 18F-FDG positron tomography and computed tomography (PET-CT). A scan was performed on nine untreated hyperthyroid patients with Graves’ disease and then again after methimazole treatment and normalization of thyroid hormone (average 77 days). None of the hyperthyroid patients with Graves’ disease had any observable active BAT prior to treatment. After 2 months of therapy, however, one of the patients did have detectable BAT. Interestingly, there was a significant decrease in 18F-FDG uptake in skeletal muscle after treatment. These results suggest that despite thyroid hormone activation of BAT in the mitochondria, there appears to be greater stimulation in other organs such as muscle, liver and heart. This suggests that in a hyperthyroid state, basal thermogenesis is increased in other targets while BAT and adaptive thermogenesis are not active.6 A case report explored BAT in an adolescent with severe primary hypothyroidism. BAT was measured by magnetic resonance imaging (MRI) and infrared thermal imaging (IRT) before and after thyroid hormone treatment. After 2 months of treatment with thyroid hormone, the subject was euthyroid with TSH decreasing from 989 μIU/mL to 4.3 μIU/mL. The subject’s supraclavicular BAT depot was significantly decreased.
These results suggest BAT may be regulated by thyroid hormone.28 Thyroid status was inversely related to BAT activity. T4 is a major suppressor of D2 activity in the BAT cell.17,29 Current discovery of BAT in adult humans has stimulated research as a potential novel target for obesity treatment and management of energy balance.13
THYROID THERMOGENESIS AND WHITE ADIPOSE TISSUE (WAT)
WAT depot can vary in genetic expression and function based on their location: subcutaneous adiposity and visceral adiposity. WAT stores excess calories in triglyceride form and some express Ucp1 and Ucp2 providing thermogenic properties via mitochondrial uncoupling. Abdominal adiposity is associated with proinflammatory cytokines and various hormonal disturbances associated with higher risk of health consequences. Thermogenesis in WAT parallels BAT induction by the sympathetic nervous system and subthermoneutral temperatures. WAT thermogenesis can be induced by spicy and high-fat diets and has shown to be protective in obesity and insulin resistance.30 In an in vitro study by Wang et al., physiologic doses of T3 were applied to determine its influence on thyroid receptor isotypes. TRα has been associated with adipogenesis, while thyroid receptor beta (TRβ) has been associated with thermogenesis. WAT cells predominantly express TRα and minimal TRβ. Findings of their research suggest that physiologic dosing of T3 decreased the expression of TRα and increased the expression of TRβ.31
THYROID THERMOGENESIS AND the HYPOTHALAMUS
Until recently thyroid metabolism was largely considered peripheral. New research in mice suggests that the CNS regulates thermogenesis by means of the autonomic nervous system (ANS), bringing to the foreground the importance of the central hypothalamic effect of thermogenesis.32 A study by Sjögren et al. suggests that central activation of thyroid hormone is independent of peripheral activation.33 Thyroid receptor TRα1 regulates hypothalamic T3 influencing sympathetic stimulation of BAT and thermogenesis.5,10–12,33 The AMP-activated protein (AMPK) receptors located in the ventral-medial aspect of the hypothalamus are inhibited by T3 resulting in a cascade of sympathetic stimulation and thermogenesis.14,34 AMPK is a key sensor and influencer of energy homeostasis.13,15,34,35 T3 inhibition of AMPK transiently induces expression of TRβ and UCP-1 contributing to thermogenesis.31,36 Animal studies demonstrate additional non-thermogenic mechanisms that can influence overall energy balance such as T3-activated orexigenic hormones such as neuro-peptide Y (NPY) and agouti-related protein (AgRP).37 NPY performs several functions, including increasing food intake and the storage of energy as fat. AgRP increases appetite and decreases metabolism and energy expenditure.
A recent study showed that obese individuals tend to have lower daytime body temperatures than normal weight individuals, and the findings suggest that normalizing low body temperature may be a useful target in the treatment of obesity.38 The discovery of BAT in adult humans has evoked an interest in researching it as a potential target for weight loss. Thyroid hormone is important in both basal and adaptive thermogenesis. Thyroid hormone increases BMR and is essential for adaptive thermogenesis.5 Thyroid hormone influences REE, a good estimate of basal thermogenesis. In order to evaluate the impact of exogenous thyroxine dosing on REE, nine medicated athyreotic humans were subject to three-dose adjustments in 6–8-week intervals. The goal was to evaluate if dose adjustments within normal range of TSH can influence REE. A significant inverse correlation between TSH and REE was demonstrated. A TSH increase of 0.1 mU/L to 10 mU/L resulted in a 15% decrease in REE.39 The variation in REE may be of clinical significance in chronically medicated hypothyroid individuals in the absence of compensatory mechanisms.
Inhibition of AMPK in the ventromedial hypothalamus via T3 is a potential pharmacologic entry point in controlling whole body energy.34 Other compounds have also been studied for their thermogenic properties, such as estradiol, berberine and those from Allium sativum (garlic). Estradiol (E2) centrally influences thermogenesis through AMPK in the hypothalamus. E2-deficient humans and mice demonstrate hyperphagia and reduced thermogenesis in BAT.40 Berberine derived from Coptis chinensis promotes UCP-1 in both BAT and WAT via AMPK inhibition in the hypothalamus. In obese db/db mice, berberine statistically improved REE, improved cold intolerance, increased production of BAT, and helped mitigate weight gain during a high-fat diet.41Allium sativum has the ability to activate AMPK peripherally in BAT as well as increase thermogenesis via adrenergic stimulation. Further, it has favourable metabolic anti-lipogenic properties such as lowering cholesterol and triglycerides levels in WAT.42
T3 is an important regulator of basal and adaptive thermogenesis in BAT, WAT, and the hypothalamus. The potential to manipulate thermogenesis is a novel means to increase energy expenditure, thereby offering a solution to the obesity epidemic and is gaining much attention. Additionally, improvements in other metabolic parameters such as adipogenesis regulation and insulin sensitivity have been demonstrated in animals. Also, inhibition of AMPK in the ventro-medial aspect of the hypothamus via T3 has demonstrated importance of this hormone in whole body energy balance. These findings are promoting further research to investigate it as another important anti-obesity target. The degree to which AMPK in the CNS influences whole body balance and whether this will be clinically relevant to human obesity is currently poorly understood and further investigations are necessary. Randomized, controlled trials in humans are needed to determine whether the use of T3 and inhibition of AMPK is a safe and effective method to promote long-term weight loss as well as to optimize metabolic status.
The author has nothing to disclose.
- Hulbert AJ.
Thyroid hormones and their effects: a new perspective.
- Freake HC,
Oppenheimer JH.
Thermogenesis and thyroid function.
- Mullur R,
Liu Y-Y,
Brent GA.
Thyroid hormone regulation of metabolism.
- Fonseca TL,
Werneck-De-Castro JP,
Castillo M, et al.
Tissue-specific inactivation of type 2 deiodinase reveals multilevel control of fatty acid oxidation by thyroid hormone in the mouse.
- Silva JE.
Thermogenic mechanisms and their hormonal regulation.
- Zhang Q,
Miao Q,
Ye H, et al.
The effects of thyroid hormones on brown adipose tissue in humans: a PET-CT study.
- Oelkrug R,
Polymeropoulos ET,
Jastroch M.
Brown adipose tissue: physiological function and evolutionary significance.
- Gardner DF,
Kaplan MM,
Stanley CA,
Utiger RD.
Effect of tri-iodothyronine replacement on the metabolic and pituitary responses to starvation.
- Müller MJ,
Bosy-Westphal A.
Adaptive thermogenesis with weight loss in humans.
- Ponrartana S,
Hu HH,
Gilsanz V.
On the relevance of brown adipose tissue in children.
- Gilsanz V,
Hu HH,
Kajimura S.
Relevance of brown adipose tissue in infancy and adolescence.
- Cannon B,
Nedergaard J.
Yes, even human brown fat is on fire! - Cannon B,
Nedergaard J.
Brown adipose tissue: function and physiological significance.
- Vijgen G,
van Marken Lichtenbelt W.
Brown adipose tissue: clinical impact of a re-discovered thermogenic organ.
- Cypess AM,
Lehman S,
Williams G, et al.
Identification and importance of brown adipose tissue in adult humans.
- Lee J-Y,
Takahashi N,
Yasubuchi M, et al.
Triiodothyronine induces UCP-1 expression and mitochondrial biogenesis in human adipocytes.
- Stephens M,
Ludgate M,
Rees DA.
Brown fat and obesity: the next big thing?
- Marrif H,
Schifman A,
Stepanyan Z, et al.
Temperature homeostasis in transgenic mice lacking thyroid hormone receptor-alpha gene products.
- Martinez-deMena R,
Anedda A,
Cadenas S,
Obregon M-J.
TSH effects on thermogenesis in rat brown adipocytes.
- Enerbäck S,
Jacobsson A,
Simpson EM, et al.
Mice lacking mitochondrial uncoupling protein are cold-sensitive but not obese.
- Ueta CB,
Olivares EL,
Bianco AC.
Responsiveness to thyroid hormone and to ambient temperature underlies differences between brown adipose tissue and skeletal muscle thermogenesis in a mouse model of diet-induced obesity.
- Stanford KI,
Middelbeek RJW,
Townsend KL, et al.
Brown adipose tissue regulates glucose homeostasis and insulin sensitivity.
- Liu X,
Zheng Z,
Zhu X, et al.
Brown adipose tissue transplantation improves whole-body energy metabolism.
- Liu X,
Wang S,
You Y, et al.
Brown adipose tissue transplantation reverses obesity in Ob/Ob mice.
- Tsao T-S,
Lodish HF,
Fruebis J.
ACRP30, a new hormone controlling fat and glucose metabolism.
- Yamauchi T,
Kamon J,
Waki H, et al.
The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity.
- Kozacz A,
Grunt P,
Steczkowska M, et al.
Thermogenic effect of glucose in hypothyroid subjects.
- Kim MS,
Hu HH,
Aggabao PC,
Geffner ME,
Gilsanz V .
Presence of brown adipose tissue in an adolescent with severe primary hypothyroidism.
- Silva JE,
Larsen PR.
Interrelationships among thyroxine, growth hormone, and the sympathetic nervous system in the regulation of 5′-iodothyronine deiodinase in rat brown adipose tissue.
- DiSilvestro D,
Petrosino J,
Aldoori A,
Melgar-Bermudez E,
Wells A,
Ziouzenkova O.
Enzymatic intracrine regulation of white adipose tissue.
- Wang C-Z,
Wei D,
Guan M-P,
Xue Y-M.
Triiodothyronine regulates distribution of thyroid hormone receptors by activating AMP-activated protein kinase in 3T3-L1 adipocytes and induces uncoupling protein-1 expression.
- López M,
Alvarez CV,
Nogueiras R,
Diéguez C.
Energy balance regulation by thyroid hormones at central level.
- Sjögren M,
Alkemade A,
Mittag J, et al.
Hypermetabolism in mice caused by the central action of an unliganded thyroid hormone receptor alpha1.
- Hardie DG.
AMPK: positive and negative regulation, and its role in whole-body energy homeostasis.
- Hardie DG,
Ross FA,
Hawley SA.
AMPK: a nutrient and energy sensor that maintains energy homeostasis.
- López M,
Varela L,
Vázquez MJ, et al.
Hypothalamic AMPK and fatty acid metabolism mediate thyroid regulation of energy balance.
- Coppola A,
Liu Z-W,
Andrews ZB, et al.
A central thermogenic-like mechanism in feeding regulation: an interplay between arcuate nucleus T3 and UCP2.
- Grimaldi D,
Provini F,
Pierangeli G, et al.
Evidence of a diurnal thermogenic handicap in obesity.
- al-Adsani H,
Hoffer LJ,
Silva JE.
Resting energy expenditure is sensitive to small dose changes in patients on chronic thyroid hormone replacement.
- Martínez de Morentin PB,
González-García I,
Martins L , et al.
Estradiol regulates brown adipose tissue thermogenesis via hypothalamic AMPK.
- Zhang Z,
Zhang H,
Li B, et al.
Berberine activates thermogenesis in white and brown adipose tissue.
- Lee M-S,
Kim I-H,
Kim C-T,
Kim Y.
Reduction of body weight by dietary garlic is associated with an increase in uncoupling protein mRNA expression and activation of AMP-activated protein kinase in diet-induced obese mice.